

This page provides a summary of recent findings from our lab. Other aspects of our research are described in the Introduction and the Current Research Projects sections. Detailed information about our research findings can be found in our Publications page.
A major focus of our research over many years has been to define the molecular mechanisms that control cell fate specification in the Arabidopsis root epidermis. In brief, we have been trying to determine what genes and molecules cause epidermal cells in the H position to always become hair cells and epidermal cells in the N position to become non-hair cells. Putting together the results from our work so far, we have come up with a model to explain how the process might work. This model is shown below:
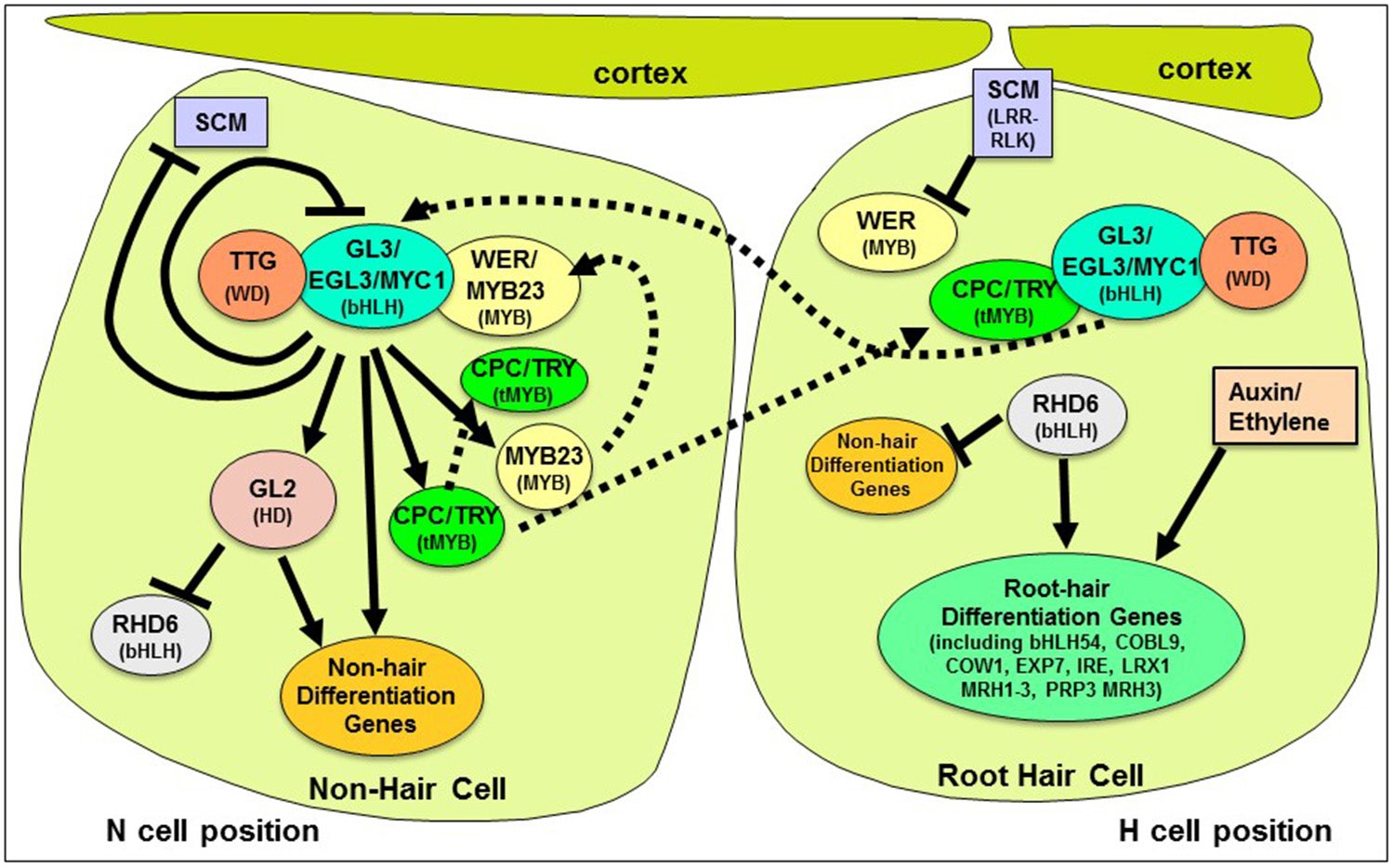 To summarize this model, the fate of an epidermal cell is determined by whether or not it expresses a genetic pathway that inhibits hair cell differentiation. If a cell does not express this inhibitory pathway, then it develops as a hair cell; if it does express this pathway, then it develops as a non-hair cell. The expression of this pathway appears to be controlled by the relative abundance of two MYB-type transcription factors, WER and CPC. Each of these is proposed to interact with the same bHLH transcription factors and with the TTG protein, but the WER protein is able to generate an active transcription complex (that causes expression of the inhibition pathway) and the CPC protein generates an inactive complex (that cannot induce expression of the the inhibition pathway. The epidermal cell pattern is proposed to result from the WER activity being relatively concentrated in the N position cells and the CPC activity being concentrated in cells in the H position.
To summarize this model, the fate of an epidermal cell is determined by whether or not it expresses a genetic pathway that inhibits hair cell differentiation. If a cell does not express this inhibitory pathway, then it develops as a hair cell; if it does express this pathway, then it develops as a non-hair cell. The expression of this pathway appears to be controlled by the relative abundance of two MYB-type transcription factors, WER and CPC. Each of these is proposed to interact with the same bHLH transcription factors and with the TTG protein, but the WER protein is able to generate an active transcription complex (that causes expression of the inhibition pathway) and the CPC protein generates an inactive complex (that cannot induce expression of the the inhibition pathway. The epidermal cell pattern is proposed to result from the WER activity being relatively concentrated in the N position cells and the CPC activity being concentrated in cells in the H position.
We have discovered several factors that influence the relative abundance of the WER and CPC. These include (1) additional functionally similar genes/proteins, including the MYB23 (similar to WER) and the TRY and ETC1 (similar to CPC), (2) multiple interlocking feedback loops that affect gene expression and/or protein accumulation in the H and N cells, and (3) the SCRAMBLED (SCM) receptor-like kinase protein, which is required for differential accumulation of the WER and CPC activities in a position-dependent manner.
Additional information about components of this model is presented below:
TRANSPARENT TESTA GLABRA (TTG) – The TTG gene encodes a protein with multiple WD repeats (Walker et al. (1999) Plant Cell 11:1337-1349). Although the protein sequence does not provide an immediate mechanistic understanding of TTG’s action, other WD-repeat proteins mediate protein-protein interactions, consistent with the view that TTG may form a complex with the bHLHs and the WER MYB to control downstream gene transcription.
Basic Helix-Loop-Helix Protein (bHLH) – bHLH transcription factors are defined by a basic/helix-loop-helix motif originally identified in the product of the proto-oncogene c-myc. Three Arabidopsis bHLH genes have a role in the root epidermis (GL3, EGL3, and MYC1). In particular, the GL3 and EGL3 play largely redundant roles in specifying the non-hair cell fate. Interestingly, there is evidence to suggest that the GL3 and EGL3 accumulate differently in the H and N cells of the developing root.
WEREWOLF (WER) – WER encodes an R2R3 Myb protein that appears to physically interact with the bHLH proteins to influence downstream gene transcription. Expression of the WER gene initiates in the earliest epidermal cell precursors (see image below), further suggesting its critical early patterning function. Notably, WER expression and protein accumulation occurs also in the H cell position, albeit at lower level than N cells.
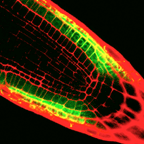
Expression of a WERpromoter-GFP transcriptional fusion in Arabidopsis roots.
GLABRA2 (GL2) – The GL2 gene encodes a homeodomain transcription factor protein (Rerie et al. (1994) Genes Dev. 8:1388-1399) that is necessary for non-hair cell differentiation. Accordingly, GL2 is preferentially expressed in the N-position cells within the meristematic and elongation regions of the root (see photos below). In contrast to ttg or wer mutations, gl2 mutations do not affect any of the other non-hair cell phenotypes (including cytoplasmic density, vacuolation, and cell division rate), suggesting GL2 only controls a subset of the non-hair cell differentiation processes.
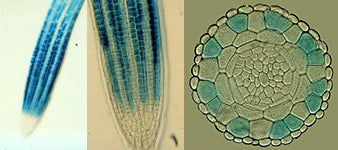
Root tip (left and center) and root cross-section (right) showing GUS activity (blue staining) in an Arabidopsis root bearing the GL2::GUS transgene. GUS expression is restricted to specific files of epidermal cells in the N position.
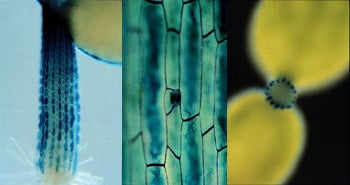
Side view (left and center) and transverse section (right) of hypocotyls from a three-day-old Arabidopsis seedlings stained for GUS activity and containing the GL2::GUS transgene. The center image shows a stomatal complex in a cell file that does not express the GL2 gene.
During the course of our work on the root epidermis, we discovered that similar cell specification mechanisms operate during epidermis development in the above-ground parts of the Arabidopsis plant. For instance, we found that the same position-dependent mechanism is used to generate a pattern of epidermal cells in the hypocotyl (the seedling stem). Although hypocotyl epidermal cells do not produce root hairs, there are two types of epidermal cells in the hypocotyl; cells that generate stomatal complexes and cells that do not. Amazingly, these two cell types arise in the same position-dependent manner as the hair and non-hair cells in the root epidermis. Stomatal cells preferentially arise outside an anticlinal cortical cell wall (analogous to the H position in the root), whereas cells located outside a periclinal cortical cell wall (analogous to the N position) preferentially develop as non-stomatal cells. Furthermore, this hypocotyl cell pattern is controlled by the same molecular components as the root pattern, including the TTG, WER, and GL2 factors. This parallel pattern of cell types and gene activities indicates that a common position-dependent cell specification mechanism is employed during development of both the root and hypocotyl epidermis of the Arabidopsis seedling.
Now that you’ve read about our past research, you can find out what we have planned for the future in the Current Research Projects section or, if you want more detailed information about this research, check out our recent publications.