But what was the time? – where was a clock? Virginia Woolf, Mrs. Dalloway
In animals the timekeeping necessary for sleep/activity rhythms takes place within neurons located in discrete regions of the central nervous system (CNS). Remarkably, neuronal circadian clocks can maintain molecular and physiological rhythms in the absence of time-cues from the environment or other cells. Despite such cell-autonomous timekeeping, the orchestration of daily behavioral rhythms depends on networks of clock neurons. Work in the Shafer lab seeks to understand the circuit properties of these networks and to examine the roles that identified neurons play in the control of circadian rhythms. We use anatomical, genetic, and live-imaging techniques in the fly Drosophila melanogaster to discover how time is kept within the brain and how it is used to orchestrate daily and seasonal changes in behavior.
The Drosophila brain consists of approximately 100,000 neurons, around 150 of which express a molecular circadian clock. The anatomy of these neurons, their targets, and their relationships with sensory inputs are one focus of our research.
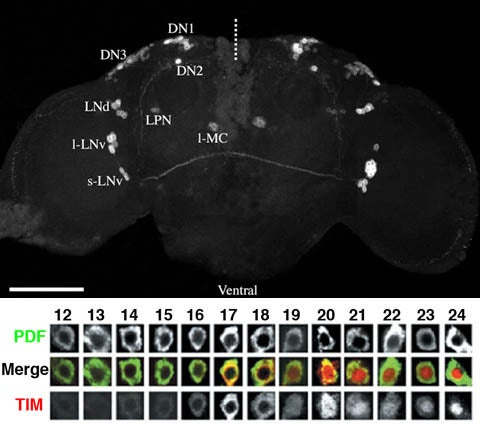
The neuroanatomy of timekeeping. Top Panel: A fly brain whose clock neurons have been visualized using an enhancer trap element called R32, which drives β-galactosidase specifically in the clock neuron network. The neurons were visualized by anti-β-galactosidase immunocytochemistry using a scanning laser confocal microscope. The various anatomical classes of clock neuron are labeled. The dashed line indicates the midline and the scale bar = 100μm. Bottom Panel: The molecular circadian rhythm in a single class of clock neuron. Each column represents an optical section through a single large ventrolateral neuron (l-vLN) double labeled for PDF, a neuropeptide, and TIMELESS (TIM), a central cog in the molecular clock. The numbers above the column indicate “Zeitgeber Time” in a 12:12 light/dark cycle (LD), where the hours from 12 to 24 indicate the dark phase of day. These neurons display a rhythm in TIM abundance and subcellular localization.
The discovery of clock genes in Drosophila changed the way we think about the genetic basis of behavior. The study of these and other behaviorally relevant genes has established that single nucleotide changes in identified genes can have significant effects on behavior, even in humans. The Shafer lab is interested in how gene function ripples through the complex neural circuitry of the brain to contribute to animal behavior. To that end we manipulate gene function within identified neurons in the fly brain to understand where clock genes are required for the normal control of behavioral rhythms. In doing so we hope to integrate the anatomical and genetic basis of timekeeping.
Exploring gene-function in specific neuron-classes in Drosophila. In the fly brain, a group of 16 neuronal clocks express a neuropeptide called PDF. The top panels show these neurons in two different flies. PDF is shown in green and PERIOD (PER), a component of the molecular clock, is shown in magenta. Scale bar = 50μm. Flies with a null mutation in PDF cannot maintain rhythms under constant darkness and temperature (DD), indicating that this neuropeptide is critical for timekeeping (Renn et al., 1999 Cell). To map where PDF is required within the clock circuitry of the fly, PDF was knocked down in specific neuronal subsets using RNA-interference. Normally PDF is express in four large and four small vLNs (l-vLNs and s-vLNs) in both brain hemispheres (top left panel). Flies with normal PDF expression maintain a bimodal activity under LD cycles and show a daily rhythm of activity under DD conditions (bottom left panel). If PDF expression is knocked down in the s-vLNs (top right panel), flies maintain a normal bimodal activity pattern under LD but cannot maintain activity rhythms under DD conditions (bottom right panel). Thus, PDF appears to be required in the s-vLNs for the maintenance of activity rhythms in the absence of environmental time-cues. By integrating anatomical and genetic analyses in this way, the Shafer lab maps the functions of behaviorally relevant genes within brain.
The considerable genetic tools available in Drosophila make it possible to manipulate gene expression and cell function with remarkable precision. However, the small nature of the fly’s CNS and the inaccessibility of most of its neurons to electrophysiological measurement make the physiological analysis of neuronal networks difficult. The use of genetically encoded, fluorescent sensors of cell signaling have made it possible to conduct physiological investigations within genetically defined networks of neurons in the fly brain. The Shafer lab uses live imaging methods to investigate the circuit properties of the fly’s clock neuron network. In addition, we are interested in developing new imaging methods for the neurophysiological investigation of neuronal networks that have been implicated in the control of various animal behaviors.
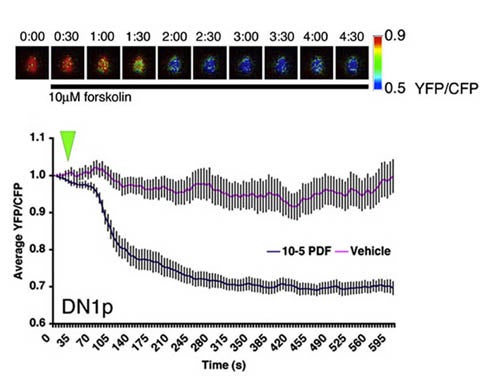
Live Imaging of Clock Neuron Networks. Neuropeptide signaling is an important component of the clock neuron network in animals. In both flies and mammals many key neuropeptides signal through G-Protein-Coupled-Receptors (GPCRs) that signal through cyclic adenosine monophosphate (cAMP). We use a genetically encoded sensor for cAMP called Epac-1-camps to observe cAMP signaling in living neurons and measure their responses to neuropeptides and other neuromodulators. The top panel shows a single l-vLN responding to forskolin, a direct activator of the adenylate cyclases that produce cAMP. The bottom panel shows the pooled responses of many clock neurons called DN1ps to PDF, a neuropeptide whose GPCR signals through cAMP. The green triangle indicates when PDF or vehicle was added to the brain.