** THIS PAGE IS CURRENTLY UNDER CONSTRUCTION **

The NSF Arctic LTER – Land-Water Interactions Research
Current Projects and Results
Introduction. The simple idea of a hydrological catchment as a study ecosystem has provided a clear framework of biogeochemical cycling within and between ecosystems for several decades. Yet it has proved an extraordinary challenge to measure the outputs of energy and biochemical elements and relate them back to the underlying processes controlling the structure and function of the terrestrial ecosystem. We still understand little about the complex dynamics and rates of production of dissolved materials on land, and their delivery to surface waters. For example, a review of 42 studies of DOC and DON concentrations and fluxes in temperate forests found that lab and field studies differed greatly in their results, and that site-specific controls such as temperature or C:N ratios were rarely evident at regional scales (Michalzik et al. 2001). We do know that across biomes the climate and vegetation influence the production of materials moving from land to water, and precipitation and hydrology are strong drivers of material exports and can govern the response of receiving surface waters. Using our conceptual model of these controls (Figure LW-1), we are asking research questions about each of them and synthesizing our observations in part by determining how these processes scale in space and time across landscapes. We have also begun to incorporate these concepts and measurements into mathematical models, necessary for extrapolations and predictions of how the arctic system operates and how it will respond to change. The sections below outline some background and highlights of our major research questions and results within the framework of land-water interactions.
In summary, the main conclusions of our research so far are: (1) Soil and especially root production of dissolved materials that can be exported to surface waters is extremely high, but the net export is low and thus processing by microbes must be substantial; (2) Microbial community composition and activity are strongly linked, and there are distinct and consistent patterns of microbial processing at key points in terrestrial and aquatic ecosystems across the landscape; (3) Interactions between different ecosystems along the toposequence of hillslopes are critical for understanding how process-level knowledge can be scaled-up to answer questions on catchment and regional biogeochemistry; and (4) Despite no evidence of increasing thaw depth near Toolik Lake, observed changes in the geochemistry of lakes and streams over time can only be explained by a melting of permafrost and thus systematic changes in thaw depth in the basin due to global warming.
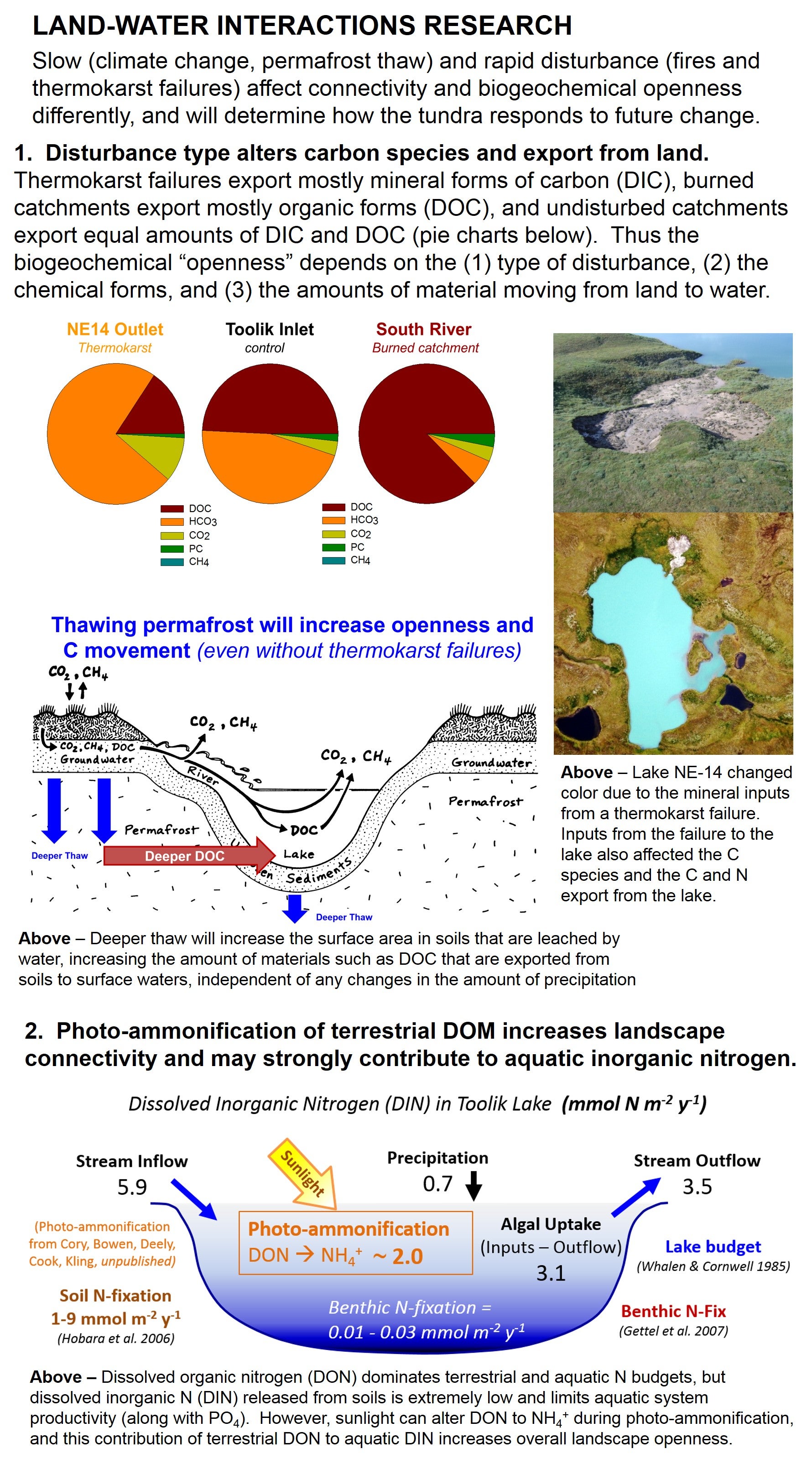
(1) Production and fate of dissolved materials on land. Our initial research in land-water interactions for the Arctic LTER focused on carbon cycling, and set the stage for current research directions. We found that the C loss from the entire Kuparuk Basin via streams and lakes is around 4 g C m-2 of land surface per year, with almost one third of this loss as CO2 and CH4 released from surface waters directly into the atmosphere (Figure LW-2). This lateral loss of C is substantial, and prior to this was unaccounted for in terrestrial budgets. Globally, this movement of gases to the atmosphere from lakes and streams is about 25% of the total global C flux from land to the oceans through rivers.
On the basis of these findings we examined in detail the production rates of dissolved C in soils, and applied a 14C tracer to intact tundra ecosystems (see Table XX). The findings were surprising in that the production of DOC from plant roots alone is extremely high, from 1-4 g C m-2 per day, whereas DOC export is only 2-3 g C m-2 per year! Our conclusion is that microbial processing of this C must be substantial, ~2 orders of magnitude higher than the net catchment export (Judd and Kling 2002; Kling, Nadelhoffer, Sommerkorn, Rastetter unpublished). And this importance of microbial processing as DOM moves from land to oceans is not restricted to the Arctic; consider that in most terrestrial systems NPP is very large, 100s of g C m-2 per year, and yet the NEP is usually near zero and dissolved export averages only ~6 g C m-2 y-1 worldwide (Hope et al. 1994). Thus the huge difference in terrestrial C production and aquatic export must be due mainly to microbial processing in soils. Given this strong biogeochemical influence across landscape scales, we can ask questions about the spatial pattern of microbial processing – where are the control points, and who is responsible.
(2) Microbial processing across the landscape. The influence of land-water transfers of nutrients and organic matter is a dominant aspect of aquatic ecology, and microbial transformations of these materials underlie production, respiration, and atmospheric gas-exchange in ecosystems. But consideration of how these processes are linked and interact across the landscape is relatively new, and requires an integration of concepts in microbial and landscape ecology. For example, we must consider the congruence of ecotones and spatial boundaries of ecosystems with the rates of microbial activity, as well as the biogeographical diversity of microbes and the time scales that microbial populations adapt physiologically and by changing population frequencies. As described below, we found several distinct patterns of microbial species and processing rates in terrestrial and aquatic ecosystems across the landscape.
Arctic ecosystems differ consistently in landscape position, plant species composition, litter biochemistry, and biogeochemical cycling rates. To test the idea that these ecosystems contain distinct microbial communities that differentially transform dissolved organic matter (DOM) as it moves downslope from dry upland to wet lowland tundra, we studied soil microbial communities in upland tussock, stream-side birch-willow, and lake-side wet sedge tundra (Figure LW-3). Using phospholipid fatty acids and 16s-rRNA analyses, we found that microbial community composition was distinct among tundra ecosystems, with tussock tundra containing a significantly greater abundance and activity of soil fungi (Judd et al. 2006; Zak and Kling 2006). We also added compound-specific 13C isotope tracers and made measurements of extracellular enzymes involved in cellulose, chitin, and lignin degradation to examine rates of microbial activity. Although the majority of 13C-labeled substrates rapidly moved into soil organic matter in all tundra soils (i.e., 50 to 90% of applied 13C), microbial respiration of labeled substrates in wet sedge tundra soil was lower than in tussock and birch-willow tundra (Figure LW-4 ; Zak & Kling 2006). Despite these differences, wet sedge tundra exhibited the greatest extracellular enzyme activity. Thus it is apparent that topographic variation in plant litter biochemistry and soil drainage shape the metabolic capability of soil microbial communities, which, in turn, influence the chemical composition of DOM across the arctic tundra landscape.
In addition to discovering these patterns of community composition and activity, we tested an ongoing debate in ecology that revolves around how species composition and ecosystem function are related. To address the mechanistic controls of this relationship, we manipulated the composition of DOM fed to aquatic bacteria to determine effects on both bacterial activity and community composition. Sites along terrestrial to aquatic flow paths were chosen to simulate movement of DOM through catchments (Figure LW-3), and DOM was fed to downslope and control bacterial communities. Bacterial production was measured and DOM chemistry and bacterial community composition (using denaturing gradient gel electrophoresis of 16S rRNA genes) were characterized following incubations. Bacterial production, DOC-specific bacterial production, and DOC consumption were greatest in mesocosms fed soil water DOM; soil water DOM enhanced lake and stream bacterial production by 320-670% relative to lake and stream controls (Judd et al. 2006). But the really novel finding was that adding upslope DOM to stream and lake bacterial communities resulted in significant changes in bacterial community composition relative to controls. In these experiments the bacterial community composition converged based on DOM source regardless of the initial inoculum (Figure LW-5). In other words, when lake bacteria were fed soil or stream DOM, the lake community assemblage shifted to resemble the species present in the soil or the stream (green and red arrows in Figure LW-5). Clearly the soil and stream bacteria were already present in the lake in undetectable numbers, but when exposed to soil or stream DOM these populations had a metabolic advantage and grew to replace the originally-dominate lake bacteria. These results demonstrate that shifts in the supply of natural DOM were followed by changes in both bacterial production and community composition, suggesting that changes in function are likely predicated on at least an initial change in the community composition. In similar experiments we also examined how photo-oxidation of DOM affected microbial activity and DOM processing along these dominant hydrological flow paths. The impacts of DOM photo-oxidation depended in part on DOM source, but also due to the relatively rapid shifts in bacterial community composition to groups better able to consume photo-products or tolerate harmful radicals (Judd et al. 2007). Overall, these results indicate that variation in DOM composition of soil and surface waters influences bacterial community dynamics, and in turn different communities control rates of carbon processing in set patterns across the landscape.
These landscape-level interactions were also observed between different aquatic ecosystems. In earlier work of the LTER, we showed that in a connected series of lakes and streams there was consistent and directional (downslope) processing of materials that produced spatial patterns in many limnological variables, and these patterns were coherent over time (Kling et al. 2000). That is, the interactions of material processing in both lakes and rivers are critical for understanding the structure and function of surface waters, especially in a landscape perspective. In recent research we have expanded these ideas to show that the processing of DOM by microbes, and the species of microbes present, vary consistently as water moves through a network of streams and lakes in the Toolik catchment. For example, in Toolik Lake itself the rate of bacterial activity was related to shifts in the source (terrestrial versus phytoplankton) and lability of DOM, and that bacterioplankton communities were composed of persistent populations present throughout the year and transient populations that appeared and disappeared (Figure LW-6; Crump et al. 2003). Shifts in community composition, measured by denaturing gradient gel electrophoresis (DGGE) of 16S rRNA genes, were associated with an annual peak in bacterial productivity driven by the large influx of labile terrestrial DOM associated with spring runoff. A second shift occurred after the terrestrial DOM flux declined and as the summer phytoplankton community developed.
Bacterioplankton community composition was also compared across 10 lakes and 14 streams within the Toolik catchment. Both lake and stream systems shared bacteria species (OTUs from DGGE analysis), and stream communities changed with distance from the upstream lake, suggesting both dispersal of species between lakes and streams as well as inoculation and dilution with bacteria from soil waters or hyporheic zones (Crump et al. 2007). At the same time, similarity in lake and stream communities shifted gradually down the catchment (Figure LW-7). We found evidence that dispersal influences bacterioplankton communities via advection and dilution (mass effects) in streams, and via inoculation and subsequent growth in lakes, and that the spatial pattern of bacterioplankton community composition was strongly influenced by interactions among soil water, stream, and lake environments. Overall these results reveal large differences in lake-specific and stream-specific bacterial community composition over restricted spatial scales (< 10 km), and suggest that geographic distance and connectivity influence the distribution of bacterioplankton communities across a landscape.
(3) Controls on biogeochemical processes and catchment export. One of the most critical issues in ecosystem research today is understanding how, exactly, do we apply our mechanistic or process-based knowledge of ecosystem function generated at small scales such as a m2 plot, to larger scales such as an entire catchment, region, or biome. There are myriad concerns and approaches related to issues of “ecological scaling”, but in our LTER we have focused on hillslopes as the “missing scale” required to transfer detailed process information to larger and larger areas (Figure LW-8). The toposequence of a hillslope represents the major ecosystem types and landscape morphology of an entire catchment, yet can be studied in depth and cohesively (e.g., Giblin et al. 1991). For example, we can monitor soil water chemistry from the hilltop to valley bottom through time, and relate the observed patterns to soil, plant, and microbial processes. The pattern of DOC concentration in soil waters on the hillslope in 2005 (Figure LW-9) shows early and late summer peaks at mid-slope, slightly elevated concentrations at the footslope near the valley floor throughout the summer, and no evidence of major transport of DOC from upslope to downslope during the summer. Our interpretation of this pattern is that most DOC production and consumption occurs in situ, which is consistent with the idea presented earlier that large amounts of DOC processing occur before DOC leaves the catchment. Preliminary data suggest that the same patterns (and interpretation) occur for other dissolved materials such as nitrogen and phosphorus, and the next step is to examine the specific processes and rates at the landscape points where concentrations are high or they change rapidly.
Although the mass of C or nutrients processed on the hillslope may be much greater than that transported downslope and into streams and lakes, the materials transported have both great impacts on the functioning of receiving surface waters, and can be substantial relative to the net C storage on land. Modeling of these landscape interactions based on a spatially linked, transect model indicates that hillslope interactions such as the downslope movement of nutrients and water may account for a 30% increase in C sequestration in tundra ecosystems over the next century (Figure LW-10; Rastetter et al. 2004).
Through monitoring and experimental manipulation, we are collecting the data needed to better model these interactions. And through this research we are developing the methodology to link plot-scale biogeochemical models with realistic hydrological models, in order to better simulate hillslope processes. In the future, we hope to scale this hill-slope model so it can be driven with coarse-scale spatial data to project our knowledge of hill-slope processes to the Pan Arctic and over the next century and beyond.
(4) Permafrost melting and biogeochemical impacts on terrestrial and aquatic ecosystems. Despite clear evidence of arctic warming, from the loss of sea ice to shifts in vegetation and species ranges, many measurements throughout the Arctic, including at Toolik Lake, show a surprising lack of permafrost melting in the soil (Figure LW-11). At the same time, we have measured substantial increases in the alkalinity of Toolik Lake since 1975, and these changes are unrelated to processes in the lake and appear instead to be caused by increased weathering of mineral soils in the catchment (Figure LW-11). This is puzzling given the fact that the depth of summer thaw has not increased over time to expose more mineral soil. However, these measurements of the maximum depth of summer thaw in soils are traditionally made using steel probes, which are limited in their use to upland terrestrial environments. To overcome this limitation we used a new approach to show that changes in the geochemistry of surface waters must be related to changes in thaw depth, although the thawing may be confined to the unfrozen zones underneath streams and lakes rather than to the entire upland catchment.
The first piece of evidence we have to build this conclusion is that the carbonate concentrations in the soils increase with depth because the deeper soils are more mineral rich, and they have been frozen so that less weathering has occurred (Keller et al. 2007). The second piece of evidence is that the ratio of strontium isotopes (87Sr/86Sr) in soils decreases with depth near the Arctic LTER (Figure LW-12). This means that as rainwater flows through deeper and deeper soils it will pick-up the soil signature and the strontium isotopic ratio in the water will decrease. Finally, we observed just such a decrease in strontium isotope ratio in the stream entering Toolik Lake over the last 10 years (Figure LW-12). The implication is that water flowpaths in the basin have progressively deepened and are now in contact with previously frozen soils of different chemistry. Because thaw depths of terrestrial sites have not changed during that time period, it is likely that the unfrozen thaw bulb found underneath streams and lakes has actually expanded, and the deeper thaw here contributes most to the altered chemistry. Our results also suggest that increasing thaw depth will lead to increasing Ca supply to soils and streams, as well as spatially variable increases in P and K supply (Keller et al. 2007). It may be that such changes in stream chemistry caused by permafrost melting are more widespread in the Arctic than currently believed, which can be tested using this new geochemical method at other sites in arctic and boreal regions with permafrost.
REFERENCES
Cole, J. J., N. Caraco, N., G. W. Kling, and T. Kratz. 1994. Carbon dioxide supersaturation in the surface waters of lakes. Science 265:1568-1570.
Crump, B. C., G. W. Kling, M. Bahr, J. E. Hobbie. 2003. Bacterioplankton community shifts in an arctic lake correlate with seasonal changes in organic matter source. Applied Environmental Microbiology 69:2253-2268.
Crump, B. C., H. E. Adams, J. E. Hobbie, G. W. Kling. In Press. Biogeography of bacterioplankton in lakes and streams of an Arctic tundra catchment. Ecology 88: .
Giblin, A.E., K.J. Nadelhoffer, G.R. Shaver, J.A. Laundre and A.J. McKerrow. 1991. Biogeochemical diversity along a riverside toposequence in arctic Alaska. Ecol. Monogr. 61:415-435.
Judd, K. E. and G. W. Kling. 2002. Production and export of dissolved C in arctic tundra mesocosms: the roles of vegetation and water flow. Biogeochemistry 60:213-234.
Judd, K.E., B.C. Crump, and G. W. Kling. 2006. Environmental drivers control ecosystem function in bacteria through changes in community composition. Ecology 87:2068-2079.
Judd, K. E., B. C. Crump, and G. W. Kling. 2007. Bacterial responses in activity and community composition to photo-oxidation of dissolved organic matter from soil and surface waters. Aquatic Sciences 69:96-107. DOI 10.1007/s00027-006-0908-4.
Keller, K., J. Blum, and G. W. Kling. 2007. Geochemistry of soils and streams on surfaces of varying ages in arctic Alaska. Arctic, Antarctic, & Alpine Research 39:84-98.
Kling, G. W. 1995. Land-water linkages: the influence of terrestrial diversity on aquatic systems, pp. 297-310. In: F. S. Chapin and C. Korner (eds.), The Role of Biodiversity in Arctic and Alpine Tundra Ecosystems, Springer-Verlag, Berlin. 320pp.
Kling, G. W., G. W. Kipphut, and M. C. Miller. 1991. Arctic lakes and rivers as gas conduits to the atmosphere: implications for tundra carbon budgets. Science 251:298-301.
Kling, G. W., G. W. Kipphut, and M. C. Miller. 1992. The flux of carbon dioxide and methane from lakes and rivers in arctic Alaska. Hydrobiologia 240:23-36.
Kling, G. W., G. W. Kipphut, M. C. Miller, and W. J. O’Brien. 2000. Integration of lakes and streams in a landscape perspective: the importance of material processing on spatial patterns and temporal coherence. Freshwater Biology 43:477-497.
Rastetter, E.B., B. L. Kwiatkowski, S. Le Dízes, and J.E. Hobbie. 2004. The Role of Down-Slope Water and Nutrient Fluxes in the Response of Arctic Hill Slopes to Climate Change. Biogeochemistry 69:37-62.
Reeburgh, W.S., J.Y. King, S.K. Regli, G.W. Kling, N.A. Auerbach, D.A. Walker. 1998. A CH4 emission estimate for the Kuparuk River Basin, Alaska. J. Geophysical Research 103:29,005-2,9013.
Zak, D. R. and G. W. Kling. 2006. Microbial Community Composition and Function across an Arctic Tundra Landscape. Ecology 87:1659-1670.